同種と異種の花粉を区別する分子を発見~種の壁を自在に制御する技術の開発に期待~
2019.07.02
- プレスリリース
- 研究
同種と異種の花粉を区別する分子を発見
~種の壁を自在に制御する技術の開発に期待~
生物は多様化することで、地球環境の変化に適応してきました。同時に、多様化は種の誕生をもたらしてきましたが、生物が自他の種を積極的に識別する分子メカニズムの存在は未知でした。
東京大学大学院農学生命科学研究科の藤井壮太助教(兼任JSTさきがけ研究者)と高山誠司教授、横浜市立大学木原生物学研究所 清水健太郎客員教授らの研究グループは、モデル植物のシロイヌナズナから異種の花粉を積極的に排除する雌しべ因子をコードする遺伝子Stigmatic Privacy 1 (SPRI1)を発見し、その機能を解析しました。SPRI1遺伝子を欠損した変異株では、通常排除されるはずの異種の花粉が侵入するようになりました。SPRI1タンパク質は雌しべの先端で花粉を受け取る部分である柱頭の細胞膜に局在して異種と自種の花粉を識別し、異種のみを排除するメカニズムに関わることを明らかにしました。SPRI1遺伝子を欠損した株では異種の花粉の侵入により正常な受精が阻害されることから、SPRI1タンパク質は異種の花粉が混在する野外環境下での種間のせめぎあいにおいて重要な役割を果たすと考えられます。種の壁を司るSPRI1タンパク質を人為的に制御することで種間交雑が容易になり、より広範な地球環境に適応する作物の開発が可能になると期待されます。本研究成果は植物生物学分野で最も権威が高いNature Plants誌に掲載されました。
本研究は、科学技術振興機構(JST) 戦略的創造研究推進事業 さきがけ「フィールドにおける植物の生命現象の制御に向けた次世代基盤技術の創出(研究総括:岡田 清孝)」における研究課題「遺伝育種の拡張に向けた種間隔離メカニズムの解明」(研究者:藤井 壮太、研究期間:2016年10月〜2020年3月)、科学研究費補助金新学術研究領域研究「植物新種誕生の原理」における課題名「初期受粉過程における種間障壁の分子基盤解明」(研究代表者:高山 誠司、研究期間:2016年7月〜2021年3月)の一環で行われました。
東京大学大学院農学生命科学研究科の藤井壮太助教(兼任JSTさきがけ研究者)と高山誠司教授、横浜市立大学木原生物学研究所 清水健太郎客員教授らの研究グループは、モデル植物のシロイヌナズナから異種の花粉を積極的に排除する雌しべ因子をコードする遺伝子Stigmatic Privacy 1 (SPRI1)を発見し、その機能を解析しました。SPRI1遺伝子を欠損した変異株では、通常排除されるはずの異種の花粉が侵入するようになりました。SPRI1タンパク質は雌しべの先端で花粉を受け取る部分である柱頭の細胞膜に局在して異種と自種の花粉を識別し、異種のみを排除するメカニズムに関わることを明らかにしました。SPRI1遺伝子を欠損した株では異種の花粉の侵入により正常な受精が阻害されることから、SPRI1タンパク質は異種の花粉が混在する野外環境下での種間のせめぎあいにおいて重要な役割を果たすと考えられます。種の壁を司るSPRI1タンパク質を人為的に制御することで種間交雑が容易になり、より広範な地球環境に適応する作物の開発が可能になると期待されます。本研究成果は植物生物学分野で最も権威が高いNature Plants誌に掲載されました。
本研究は、科学技術振興機構(JST) 戦略的創造研究推進事業 さきがけ「フィールドにおける植物の生命現象の制御に向けた次世代基盤技術の創出(研究総括:岡田 清孝)」における研究課題「遺伝育種の拡張に向けた種間隔離メカニズムの解明」(研究者:藤井 壮太、研究期間:2016年10月〜2020年3月)、科学研究費補助金新学術研究領域研究「植物新種誕生の原理」における課題名「初期受粉過程における種間障壁の分子基盤解明」(研究代表者:高山 誠司、研究期間:2016年7月〜2021年3月)の一環で行われました。
発表者
藤井 壮太 (東京大学大学院農学生命科学研究科応用生命化学専攻 助教
/科学技術振興機構(JST)さきがけ研究者)
土松 隆志 (千葉大学大学院理学研究院生物学研究部門 准教授)
木村 友香 (東京大学大学院農学生命科学研究科応用生命化学専攻 技術職員)
石田 翔太 (東京大学大学院農学生命科学研究科応用生命化学専攻 修士課程2年)
下里 裕子 (奈良先端科学技術大学院大学バイオサイエンス研究科 博士研究員:当時)
岩野 恵 (奈良先端科学技術大学院大学バイオサイエンス研究科 助教:当時)
古川 翔子 (奈良先端科学技術大学院大学バイオサイエンス研究科 博士前期課程:当時)
糸山 和香 (奈良先端科学技術大学院大学バイオサイエンス研究科 博士前期課程:当時)
和田七夕子 (奈良先端科学技術大学院大学先端科学技術研究科 助教)
清水健太郎 (University of Zurich, Department of Evolutionary Biology and Environmental Studies 教授
/横浜市立大学木原生物学研究所 客員教授)
高山 誠司 (東京大学大学院農学生命科学研究科応用生命化学専攻 教授)
藤井 壮太 (東京大学大学院農学生命科学研究科応用生命化学専攻 助教
/科学技術振興機構(JST)さきがけ研究者)
土松 隆志 (千葉大学大学院理学研究院生物学研究部門 准教授)
木村 友香 (東京大学大学院農学生命科学研究科応用生命化学専攻 技術職員)
石田 翔太 (東京大学大学院農学生命科学研究科応用生命化学専攻 修士課程2年)
下里 裕子 (奈良先端科学技術大学院大学バイオサイエンス研究科 博士研究員:当時)
岩野 恵 (奈良先端科学技術大学院大学バイオサイエンス研究科 助教:当時)
古川 翔子 (奈良先端科学技術大学院大学バイオサイエンス研究科 博士前期課程:当時)
糸山 和香 (奈良先端科学技術大学院大学バイオサイエンス研究科 博士前期課程:当時)
和田七夕子 (奈良先端科学技術大学院大学先端科学技術研究科 助教)
清水健太郎 (University of Zurich, Department of Evolutionary Biology and Environmental Studies 教授
/横浜市立大学木原生物学研究所 客員教授)
高山 誠司 (東京大学大学院農学生命科学研究科応用生命化学専攻 教授)
発表のポイント
|
発表内容
種と種の間には生殖障壁(注1)があります。特に精細胞と卵細胞が受精する前に起こる種間の不和合性は受精前障壁と呼ばれており(図a)、有限の資源を好ましくない子孫に分配することを避けるメカニズムであると考えられてきました。しかし、受精前障壁のメカニズムについてはほとんど未解明でした。
本研究グループはモデル植物であるシロイヌナズナ(Arabidopsis thaliana)とアブラナ科の近縁植物種を用いて異種間の交雑試験を行い、同種と異種の花粉を識別するメカニズムの解明を目指しました。多くの系統間で種間交雑実験を試したところ、マルコルミア・リットレア(Malcolmia littorea)というアブラナ科植物種(図b)の花粉がシロイヌナズナのある系統(Col-0)では排除されるのに対し、別の系統(Cvi-0)では雌しべ内に侵入することを発見しました。そこでシロイヌナズナの338種類の野生系統の全ゲノム配列情報を利用し、ゲノムワイド関連解析(注2)という統計手法を用いてM. littoreaの花粉の排除能力を決定する原因遺伝子座(注3)を探索しました。その結果、第4四染色体上の単一の遺伝子座が排除能力に大きく寄与することを見いだしました。この染色体領域内で雌しべにおいて発現する候補原因遺伝子を破壊した系統では、野生型のCol-0では排除されるはずのM. littoreaの花粉が、雌しべの内部にまで侵入することが明らかになりました(図c)。この原因遺伝子は雌しべの先端で花粉を受け取る部分である柱頭(Stigma)で「種」の壁として機能すると考えられ、この機能は他者からの侵害を受けない権利(プライバシー)に通じることから、この原因遺伝子をStigmatic Privacy 1(SPRI1)と命名しました。SPRI1タンパク質は、雌しべの柱頭でのみ機能し(図d)、細胞膜を4回貫通する領域を持つことが示唆されました。
半数近くの植物種は、同種内でも自己の花粉とは受精せず、非自己のみと受精して子孫を残す自家不和合性(注4)という性質を持っていることが知られています。自家不和合種の雌しべは自己の花粉で受精する自家和合種の花粉は受け入れませんが、自家和合種は自家不和合種の花粉を受け入れる「種の一側性不和合性(注5)」という現象が1940年代から知られていました。このことは自家和合種においては異種の花粉を排除する能力が失われる傾向にあることを意味していますが、それを説明できる分子は明らかにされていませんでした。本研究ではDNA配列解析によりシロイヌナズナの進化の過程で少なくとも6回SPRI1遺伝子の機能が失われたことを明らかにしました。これはシロイヌナズナが自家和合性の獲得によってSPRI1の機能を維持する理由がなくなったことに起因すると考えられました。本研究によりこれまで合理的な説明がなされてこなかった種間の一側性不和合性を分子レベルで説明できるようになりました。また、ゲノム編集法を用いた解析により、SPRI1タンパク質は自家不和合性を引き起こす分子メカニズムとは完全に独立した働きを持つことも示しました。
さらにSPRI1タンパク質はM. littoreaのみならず、多様な種の花粉の排除に関わることを明らかにしました。SPRI1タンパク質の機能を破壊した系統に、自種の花粉を受粉させるより前に異種の花粉を受粉しておくと著しく受精効率が下がることが明らかとなりました。動けない植物は昆虫や風などの媒介によって受粉するため、雌しべには様々な花粉が運搬されてくる可能性があります。SPRI1タンパク質は異種の花粉が混在する野外環境下での種間のせめぎあいにおいて重要な役割を果たしていると考えられます。
精細胞と卵細胞の受精に関わる因子として哺乳類ではZP2、CD9、JUNO-IZUMO1、植物ではGCS1、LURE1-PRK6などが知られています。同種間ではこれらの雌雄タンパク質の相性が適合していることで受精が成功し、異種間では相性が悪いため受精が失敗することが報告されています。一方で、雌しべが好ましくない花粉を積極的に排除するのがSPRI1タンパク質の働きです。本研究によって、これまで知られているものとはまったく異なる分子メカニズムで配偶子を選択する仕組みを植物(シロイヌナズナ)が備えていることが初めて明らかになりました。
近年、農業への期待は多様化しており、収量、品質、機能性などの向上や変動する地球環境に適応した作物の開発が求められています。種の壁を司るSPRI1遺伝子をゲノム編集によって人為的に改変したり、SPRI1タンパク質の機能を特異的に阻害する化合物を用いて制御したりすることにより、これまでは種の壁の制約により利用が困難であった遺伝資源にも活用の道が開かれ、育種の効率が向上するとともに、新たな機能を備えた有用な作物の開発が加速すると期待されます。
本研究グループはモデル植物であるシロイヌナズナ(Arabidopsis thaliana)とアブラナ科の近縁植物種を用いて異種間の交雑試験を行い、同種と異種の花粉を識別するメカニズムの解明を目指しました。多くの系統間で種間交雑実験を試したところ、マルコルミア・リットレア(Malcolmia littorea)というアブラナ科植物種(図b)の花粉がシロイヌナズナのある系統(Col-0)では排除されるのに対し、別の系統(Cvi-0)では雌しべ内に侵入することを発見しました。そこでシロイヌナズナの338種類の野生系統の全ゲノム配列情報を利用し、ゲノムワイド関連解析(注2)という統計手法を用いてM. littoreaの花粉の排除能力を決定する原因遺伝子座(注3)を探索しました。その結果、第4四染色体上の単一の遺伝子座が排除能力に大きく寄与することを見いだしました。この染色体領域内で雌しべにおいて発現する候補原因遺伝子を破壊した系統では、野生型のCol-0では排除されるはずのM. littoreaの花粉が、雌しべの内部にまで侵入することが明らかになりました(図c)。この原因遺伝子は雌しべの先端で花粉を受け取る部分である柱頭(Stigma)で「種」の壁として機能すると考えられ、この機能は他者からの侵害を受けない権利(プライバシー)に通じることから、この原因遺伝子をStigmatic Privacy 1(SPRI1)と命名しました。SPRI1タンパク質は、雌しべの柱頭でのみ機能し(図d)、細胞膜を4回貫通する領域を持つことが示唆されました。
半数近くの植物種は、同種内でも自己の花粉とは受精せず、非自己のみと受精して子孫を残す自家不和合性(注4)という性質を持っていることが知られています。自家不和合種の雌しべは自己の花粉で受精する自家和合種の花粉は受け入れませんが、自家和合種は自家不和合種の花粉を受け入れる「種の一側性不和合性(注5)」という現象が1940年代から知られていました。このことは自家和合種においては異種の花粉を排除する能力が失われる傾向にあることを意味していますが、それを説明できる分子は明らかにされていませんでした。本研究ではDNA配列解析によりシロイヌナズナの進化の過程で少なくとも6回SPRI1遺伝子の機能が失われたことを明らかにしました。これはシロイヌナズナが自家和合性の獲得によってSPRI1の機能を維持する理由がなくなったことに起因すると考えられました。本研究によりこれまで合理的な説明がなされてこなかった種間の一側性不和合性を分子レベルで説明できるようになりました。また、ゲノム編集法を用いた解析により、SPRI1タンパク質は自家不和合性を引き起こす分子メカニズムとは完全に独立した働きを持つことも示しました。
さらにSPRI1タンパク質はM. littoreaのみならず、多様な種の花粉の排除に関わることを明らかにしました。SPRI1タンパク質の機能を破壊した系統に、自種の花粉を受粉させるより前に異種の花粉を受粉しておくと著しく受精効率が下がることが明らかとなりました。動けない植物は昆虫や風などの媒介によって受粉するため、雌しべには様々な花粉が運搬されてくる可能性があります。SPRI1タンパク質は異種の花粉が混在する野外環境下での種間のせめぎあいにおいて重要な役割を果たしていると考えられます。
精細胞と卵細胞の受精に関わる因子として哺乳類ではZP2、CD9、JUNO-IZUMO1、植物ではGCS1、LURE1-PRK6などが知られています。同種間ではこれらの雌雄タンパク質の相性が適合していることで受精が成功し、異種間では相性が悪いため受精が失敗することが報告されています。一方で、雌しべが好ましくない花粉を積極的に排除するのがSPRI1タンパク質の働きです。本研究によって、これまで知られているものとはまったく異なる分子メカニズムで配偶子を選択する仕組みを植物(シロイヌナズナ)が備えていることが初めて明らかになりました。
近年、農業への期待は多様化しており、収量、品質、機能性などの向上や変動する地球環境に適応した作物の開発が求められています。種の壁を司るSPRI1遺伝子をゲノム編集によって人為的に改変したり、SPRI1タンパク質の機能を特異的に阻害する化合物を用いて制御したりすることにより、これまでは種の壁の制約により利用が困難であった遺伝資源にも活用の道が開かれ、育種の効率が向上するとともに、新たな機能を備えた有用な作物の開発が加速すると期待されます。
発表雑誌
雑誌名:「Nature Plants」(7月1日オンライン版)
論文タイトル:Identification of a stigmatic gene functions in inter-species incompatibility in the Brassicaceae
著者:Sota Fujii*, Takashi Tsuchimatsu, Yuka Kimura, Shota Ishida, Surachat Tangpranomkorn, Hiroko Shimosato-Asano, Megumi Iwano, Shoko Furukawa, Wakana Itoyama, Yuko Wada, Kentaro K. Shimizu, Seiji Takayama*
DOI番号:10.1038/s41477-019-0444-6
アブストラクトURL:https://www.nature.com/articles/s41477-019-0444-6
論文タイトル:Identification of a stigmatic gene functions in inter-species incompatibility in the Brassicaceae
著者:Sota Fujii*, Takashi Tsuchimatsu, Yuka Kimura, Shota Ishida, Surachat Tangpranomkorn, Hiroko Shimosato-Asano, Megumi Iwano, Shoko Furukawa, Wakana Itoyama, Yuko Wada, Kentaro K. Shimizu, Seiji Takayama*
DOI番号:10.1038/s41477-019-0444-6
アブストラクトURL:https://www.nature.com/articles/s41477-019-0444-6
用語解説
(注1)生殖障壁
異なる種の間で、雌雄の配偶子の間に不適合があり、次世代を残すことができなくなるメカニズムの総称。
(注2)ゲノムワイド関連解析
ある集団において、個体間の形質の違いとDNA配列の違いとの関わりを全ゲノム配列にわたって統計的に検出する解析手法。
(注3)原因遺伝子座
染色体上で、個体間の形質の違いの原因となる遺伝子が座乗する部位。
(注4)自家不和合性
植物が自己の花粉と集団内の非自己の花粉を識別して、非自己のみと受精し子孫を作る性質。近親交配による有害な遺伝子の集積を回避し、集団内の遺伝的多様性を保つ効果があると考えられている。アブラナ科ではSP11という花粉タンパク質とSRKという雌しべタンパク質が自分自身のタイプのみと直接的に相互作用することで自己拒絶反応が起こる。
(注5)種の一側性不和合性
自家不和合性種の雌しべは自家和合性種の花粉を排除するが、その逆は受け入れられるという現象。この現象はアブラナ科を含む広範な植物種で見られることが報告されている。
異なる種の間で、雌雄の配偶子の間に不適合があり、次世代を残すことができなくなるメカニズムの総称。
(注2)ゲノムワイド関連解析
ある集団において、個体間の形質の違いとDNA配列の違いとの関わりを全ゲノム配列にわたって統計的に検出する解析手法。
(注3)原因遺伝子座
染色体上で、個体間の形質の違いの原因となる遺伝子が座乗する部位。
(注4)自家不和合性
植物が自己の花粉と集団内の非自己の花粉を識別して、非自己のみと受精し子孫を作る性質。近親交配による有害な遺伝子の集積を回避し、集団内の遺伝的多様性を保つ効果があると考えられている。アブラナ科ではSP11という花粉タンパク質とSRKという雌しべタンパク質が自分自身のタイプのみと直接的に相互作用することで自己拒絶反応が起こる。
(注5)種の一側性不和合性
自家不和合性種の雌しべは自家和合性種の花粉を排除するが、その逆は受け入れられるという現象。この現象はアブラナ科を含む広範な植物種で見られることが報告されている。
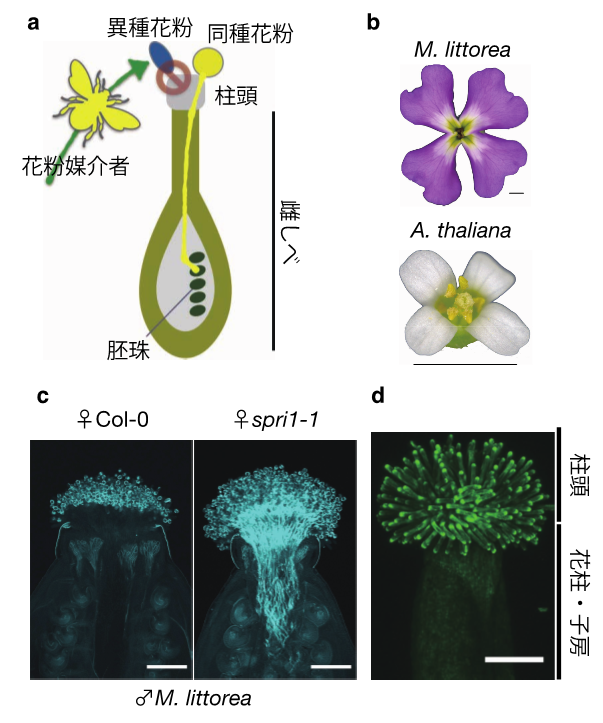
図 a) 植物では一般的に異種の花粉が排除され、同種の花粉のみが受け入れられる。b) 本研究で用いたM. littoreaとA. thalianaの花器官の外観。スケールバー:2 mm。c) 雌しべ中の花粉管を細胞壁染色試薬アニリンブルーで染色し共焦点レーザー顕微鏡で観察した。野生型(Col-0)の雌しべではM. littoreaの花粉管の侵入が見られない。一方spri1の変異体では多数の花粉管侵入が観察された。スケールバー:200 µm。d) SPRI1遺伝子の発現部位の観察。SPRI1プロモーターに蛍光タンパク質(Venus)遺伝子を連結しシロイヌナズナに導入したところ、花粉を受け取る部分である雌しべ先端の柱頭でのみ蛍光が観察された。スケールバー:200 µm。
問い合わせ先
(研究の内容に関するお問い合わせ)
横浜市立大学木原生物学研究所 客員教授 清水健太郎
(スイス・チューリッヒ大学 進化生物・環境学研究所 兼任)
TEL:+41 44 63 56740(チューリッヒ大学)
TEL:045-820-2429(木原生物学研究所)
E-mail:kentaro.shimizu@ieu.uzh.ch
(取材対応窓口、資料請求など)
研究企画・産学連携推進課 研究企画担当
Tel:045-787-2527
E-mail:kenkyupr@yokohama-cu.ac.jp